La mitocondria es el único orgánulo que el cuerpo fabrica de nuevo cuando entrenas. Pero el tipo de entrenamiento dicta si construyes contenido, función o nada.
Volumen, intensidad y modalidad hacen cosas distintas, y a veces opuestas. La divulgación ha mezclado las tres y ha vendido la zona 2 como si fuera la respuesta completa para fabricar mitocondrias.
No lo es. Cuando bajas a la biopsia muscular humana, que es el único dominio que zanja la discusión, el cuadro es más incomodo para el eslogan: la zona 2 aislada se queda corta en quien tiene horas limitadas, y la fuerza, por sí sola, no fabrica una sola mitocondria nueva.
Esta pieza es el satélite de entrenamiento del concepto de biogénesis mitocondrial: si quieres el qué es y el mecanismo molecular, esa es la entrada de referencia. Aquí bajamos a lo accionable, qué tipo de ejercicio, con qué volumen y a qué intensidad, comparado en el único terreno que importa: lo que se ve en la biopsia.
El hub de la mitocondria estableció que el ejercicio es la única intervención con evidencia humana sólida para inducir biogénesis. Correcto, pero demasiado ancho para accionarlo. Este artículo hace tres cosas: distingue las tres dimensiones de la biogénesis, compara HIIT, continuo moderado y fuerza en biopsia, y deriva una prescripción semanal que respeta la evidencia y descarta el folclore.
Contenido, función y red: tres cosas que se confunden
El término «biogénesis mitocondrial» se usa en divulgación como si apuntara a una sola cosa, fabricar mitocondrias nuevas. La definición del proceso la damos en el pilar del concepto; lo que importa aquí, para interpretar bien la evidencia del entrenamiento, es que la literatura primaria distingue al menos tres dimensiones que no siempre se correlacionan. Confundirlas es la fuente de la mitad de las afirmaciones infladas.
Contenido
El contenido mitocondrial es la cantidad total de masa mitocondrial en el músculo. Se mide por actividad de la citrato sintasa (una enzima del ciclo de Krebs usada como proxy fiable), por densidad de volumen mitocondrial en microscopía electrónica o por abundancia de proteínas de la cadena respiratoria. Aumentar contenido es añadir más fábricas de ATP.
Función respiratoria
La función respiratoria es la eficiencia intrínseca de cada unidad mitocondrial: cuánto oxígeno consume, cuánto ATP sintetiza, con qué acoplamiento. Se mide por respirometría de alta resolución (sistema Oroboros) sobre fibras musculares permeabilizadas. Puedes tener más mitocondrias pero peores, o menos pero mejores. No es lo mismo que contenido, y esta distinción va a ordenar todo lo que sigue.
Red mitocondrial
La red mitocondrial es cómo las unidades individuales se organizan en redes conectadas mediante fusión (MFN1, MFN2, OPA1) y se separan por fisión (DRP1, FIS1). La dinámica de la red determina si las mitocondrias pueden compartir contenido, reparar daños y eliminar fracciones disfuncionales por mitofagia. Una red bien regulada importa tanto o más que el número absoluto.
Esta distinción no es pedantería académica, es operativa: distintos tipos de ejercicio mueven estas variables de forma distinta y a veces en direcciones opuestas. Un trabajo reciente que resume bien el campo (Memme, Sanfrancesco y Hood, en Annual Review of Physiology de 2025) lo plantea así: pensar en biogénesis como variable única es un error categorial que explica por qué estudios con diseños similares obtienen resultados contradictorios [1].
Los sensores del ejercicio que encienden PGC-1α
El regulador maestro de la biogénesis es PGC-1α, y su cascada completa (NRF1, NRF2, TFAM y la coordinación de los dos genomas) la detallamos en el pilar. Lo relevante para el entrenamiento es qué lo activa, porque ahí está la clave de por qué unas modalidades funcionan y otras no.
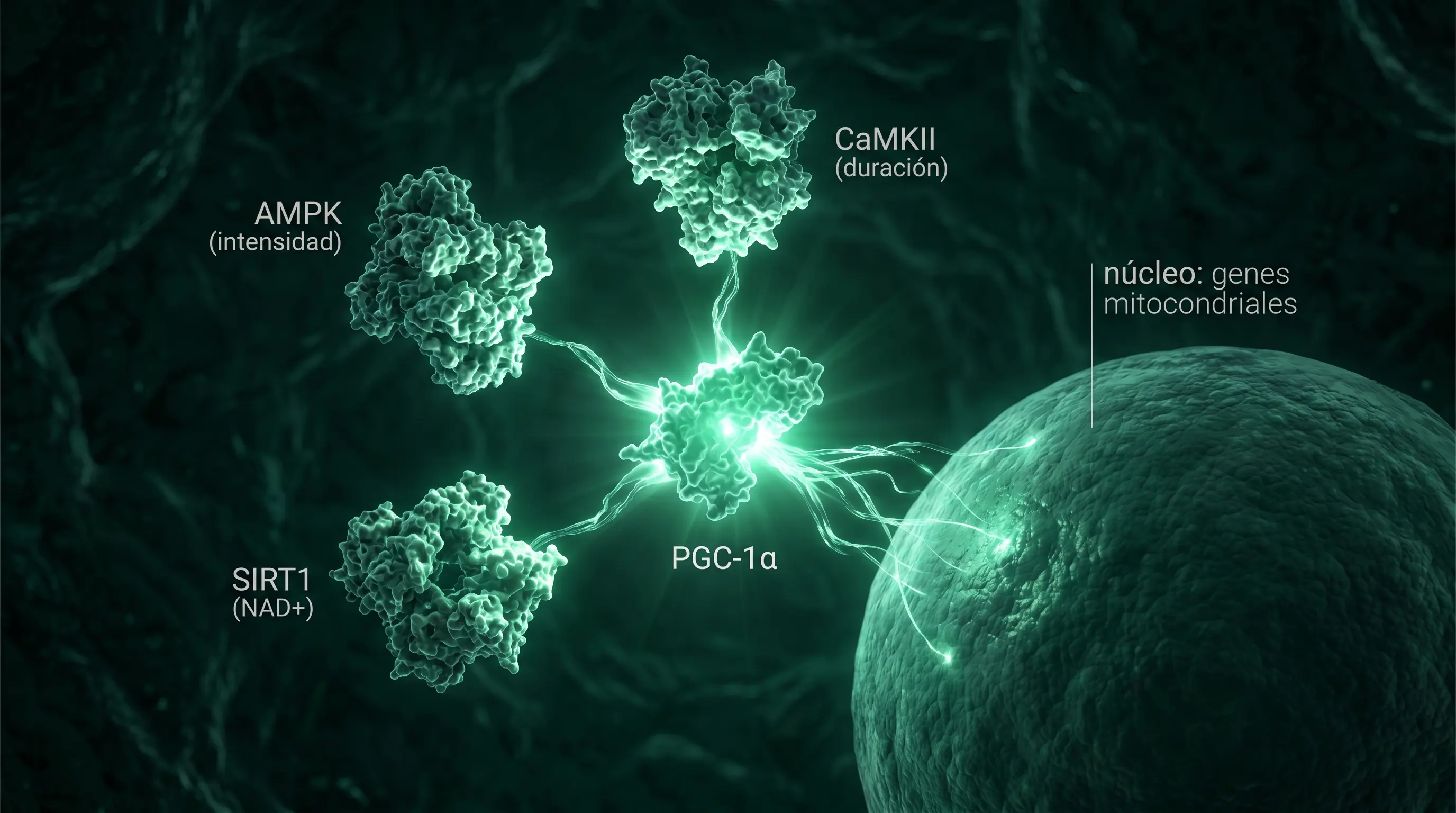
PGC-1α en una frase
Lo identificaron Puigserver y Spiegelman en 1998 sobre tejido adiposo pardo [2] y un año después Wu y Spiegelman confirmaron su papel en músculo [3]. Es el director de orquesta que coordina el genoma nuclear y el mitocondrial. Cuando se activa, la maquinaria mitocondrial se expande. Para el entrenamiento, lo que cuenta es la cadena de sensores que está aguas arriba.
AMPK, CaMKII y SIRT1: intensidad, duración y redox
Hay dos sensores principales. La AMPK se activa cuando baja la relación ATP/AMP, es decir, cuando la célula gasta energía más rápido de la que repone; fosforila directamente a PGC-1α, y su señal es proporcional a la intensidad: cuanto más duro el ejercicio, más se activa. La CaMKII se activa por las oscilaciones de calcio de la contracción y es teóricamente sensible a la duración más que a la intensidad, por eso se postula como la vía del trabajo prolongado de bajo grado; pero la evidencia es más débil de lo que el eslogan sugiere, y revisiones recientes (Gurd et al. 2023) observan respuestas inconsistentes de la CaMKII a intensidades de zona 2 [4]. Participa además SIRT1, sensor del ratio NAD+/NADH, que desacetila PGC-1α y liga la biogénesis al estado redox y al NAD disponible (la razón por la que se han estudiado precursores como NR y NMN, con resultados humanos modestos).
La biología no premia el tiempo sin perturbación. La señalización que dispara biogénesis es proporcional al estrés metabólico que el ejercicio genera, no a cuántos minutos hagas. Tres horas de paseo a 60 % de la frecuencia cardiaca máxima pueden activar PGC-1α menos que 20 minutos de intervalos por encima del umbral láctico.
El pulso oxidativo es señal, no ruido
Un matiz con consecuencias prácticas: Ristow y colaboradores demostraron en 2009, en PNAS, que suplementar dosis altas de vitamina C y E durante el entrenamiento bloquea la activación de PGC-1α y de las enzimas antioxidantes endógenas, anulando la mejora de sensibilidad a la insulina [5]. El pulso oxidativo posterior al ejercicio es la señal que queremos activar; secuestrarlo con antioxidantes exógenos es interferir con la vía. Más antioxidante no es más longevidad.
HIIT vs continuo moderado vs fuerza: lo que muestra el músculo
Toda la teoría anterior se resuelve en un único tribunal: la biopsia muscular humana con respirometría. Y ahí el cuadro es nítido cuando se separa el contenido de la función.
El estudio que lo zanja: Robinson 2017
El trabajo más limpio sobre qué ejercicio induce qué adaptación es Robinson, Konopka y Nair, en Cell Metabolism (2017, Mayo Clinic) [6]. Diseño sin interpretaciones ambiguas: 72 participantes jóvenes (18-30) y mayores (65-80), aleatorizados a 12 semanas de HIIT, entrenamiento de fuerza o entrenamiento combinado, con biopsia de vasto lateral, respirometría de alta resolución, transcriptómica y proteómica. El resultado crítico: solo el HIIT y el combinado mejoraron la capacidad aeróbica (el VO2max) y la respiración mitocondrial. La fuerza sola mejoró sensibilidad a la insulina y masa magra (adaptaciones valiosas, pero no mitocondriales intrínsecas). El HIIT fue el que más revirtió el proteóma asociado a la edad.
HIIT por encima del continuo moderado: Pengam 2025
Un estudio independiente de 2025 en Frontiers in Physiology (Pengam y colaboradores) comparó HIIT frente a entrenamiento continuo de intensidad moderada durante 6 semanas, con biopsia antes, a las 24 horas y al final [7]. Ambos aumentaron VO2max, citrato sintasa y actividad del complejo I, pero la conclusión explícita de los autores fue que el HIIT mostró efectividad superior en biogénesis y densidad de volumen mitocondrial. En cifras de apoyo, la respiración mitocondrial sube un ~22 % ya tras 2 semanas de HIIT en varones no entrenados (Jacobs 2013), y la citrato sintasa aumenta un 40-50 % tras 3 semanas de alto volumen (Granata 2016); el umbral mínimo aproximado para inducir adaptaciones ronda el 65 % de la potencia máxima.
Lo que no hace la fuerza aislada
El hallazgo de Robinson merece énfasis porque va contra una línea de marketing creciente que presenta la fuerza como «rehabilitadora mitocondrial» en sí misma. La fuerza es imprescindible por otras razones (masa muscular, densidad ósea, sensibilidad a la insulina vía GLUT4), pero no induce biogénesis comparable al aeróbico de alta intensidad. El mecanismo es coherente: la fuerza activa más mTORC1 (síntesis proteica e hipertrofia) que AMPK (biogénesis). El corolario es que no se sustituyen, son complementarios, lo que alinea con la tríada de VO2max, fuerza y masa muscular.
El matiz de la metformina
Todo lo anterior está condicionado a un factor importante. Konopka y colaboradores publicaron en 2019, en Aging Cell, un ensayo doble ciego con 53 adultos mayores que recibieron placebo o metformina durante 12 semanas de HIIT [8]. La metformina anuló el aumento de VO2max y bloqueó la mejora de respiración mitocondrial inducida por el entrenamiento. En no diabéticos que entrenan, prescribir metformina «por longevidad» es contraproducente para exactamente la adaptación que el ejercicio intenta construir.
Volumen o intensidad, y por qué la pregunta está mal formulada
En la última década, la fisiología del ejercicio ha vivido una controversia que algunos autores bautizaron como the mitochondrial content contest. Dos campos bien definidos, y la sensación de que se contradicen. No se contradicen: miden cosas distintas.
Los dos campos
Por un lado, Granata, Bishop y colegas (Victoria University): un metaanálisis que agrupa más de cincuenta estudios no encontró asociación significativa entre la intensidad relativa y los cambios en contenido mitocondrial (citrato sintasa o densidad de volumen), pero sí una relación positiva con el volumen total. Su conclusión: el volumen es el motor del contenido [9][10]. Por otro, MacInnis, Gibala y colegas (McMaster University): igualando la carga total de trabajo, la intensidad marca diferencias, especialmente para la función mitocondrial y para la señalización molecular (PGC-1α, p53) [11].
La síntesis: dos ejes, no uno
¿Quién tiene razón? Ambos, porque miden dimensiones distintas. La síntesis que ha emergido en 2023-2025 es limpia: el volumen construye contenido; la intensidad construye función. Para aumentar la masa mitocondrial total importa el tiempo acumulado de trabajo; para mejorar cómo respira cada mitocondria importa la intensidad del estímulo. No son sustituibles, son dos ejes de la misma adaptación, y optimizar uno solo es optimizar media foto. El propio Granata lo documentó: en 2016 mostró que la intensidad modula PGC-1α, p53 y respiración pero no los marcadores de contenido [10], y que el volumen determina el contenido y que las adaptaciones revierten rápido cuando el volumen cae [12].
El problema editorial de la zona 2
Aquí entra la pieza que confronta la narrativa dominante. Storoschuk, Moran-MacDonald, Gibala y Gurd publicaron en junio de 2025, en Sports Medicine, una revisión con título explícito: «Much Ado About Zone 2» [13]. Revisa las dos afirmaciones fundacionales del marketing de la zona 2 para población general (que es la intensidad óptima para mejorar la capacidad mitocondrial y la oxidación de grasas) y concluye que la evidencia no las sostiene. La zona 2 induce cambios modestos o nulos en el sensor aguas arriba de la AMPK; la señal de PGC-1α tras zona 2 es inconsistente y solo aparece con sesiones de 60-90 minutos, no de 30; la vía de la CaMKII es escasa y contradictoria. Para mejorar la capacidad mitocondrial en personas insuficientemente activas, la evidencia apoya entrenar deliberadamente por encima de la zona 2.
Un apunte de transparencia obligado: Martin J. Gibala, coautor del trabajo, declara equity en Longevity League Ltd., una empresa cuyo modelo comercial incluye prescripción de HIIT, y la declaración consta en el propio paper. No invalida los argumentos (la revisión está sometida a peer review), pero exige lectura crítica en un texto que confronta la narrativa de la zona 2. La carencia de fondo (faltan ensayos humanos que comparen zona 2 aislada frente a HIIT igualando volumen para endpoints mitocondriales) está bien documentada con independencia del conflicto de interés.
Cómo se traduce esto a una semana real
Los atletas de endurance de élite entrenan 20-30 horas semanales con distribución polarizada 80/20. Ese modelo funciona porque el volumen absoluto es enorme y ese 80 % de trabajo aeróbico moderado ya aporta señal suficiente. Extrapolarlo al adulto que entrena 3-5 horas a la semana no está justificado: a esos volúmenes, dedicar el 80 % del tiempo a zona 2 es renunciar al estímulo intenso que mueve la aguja.
| Modalidad | Activación molecular | Contenido | Función | Ventana |
|---|---|---|---|---|
| Zona 2 aislada (< umbral) | Modesta. Requiere sesiones >60-90 min | Solo con volumen alto | Evidencia limitada | Semanas-meses |
| Continuo por encima del umbral | Consistente | Sí | Sí | Semanas |
| HIIT (80-95 % Wmax) | Robusta. AMPK + CaMKII + p53 | Sí, sobre todo con volumen | Sí, claramente superior | 2-4 semanas |
| SIT (sprint, all-out) | Máxima en poco tiempo | Sí, efecto agudo grande | Sí | Alta eficiencia |
| Fuerza aislada | mTOR >> AMPK | No | No (Robinson 2017) | N/A mitocondria |
| Combinado (HIIT + fuerza) | Completa | Sí | Sí | Semanas |
La tabla no invalida la zona 2 como herramienta. La zona 2 tiene valor real (coste de recuperación bajo, bajo riesgo de lesión, sostenible día tras día), lo que la hace el vehículo ideal para acumular volumen. Lo que la evidencia no sostiene es que la zona 2 sola sea la respuesta óptima para construir mitocondrias en quien tiene horas limitadas.
Una semana orientada a salud mitocondrial
Para el adulto no atleta que busca salud mitocondrial y longevidad, una prescripción que respeta la literatura humana se ve así.
| Día | Tipo | Duración | Intensidad | Qué construye |
|---|---|---|---|---|
| Lunes | HIIT o umbral | 30-40 min | 4×4 min al 85-90 % FCmax | Función, VO2max, PGC-1α |
| Martes | Fuerza compuesta | 45-60 min | 3-5 series, 6-10 reps, 75-85 % 1RM | Masa muscular, hueso, GLUT4 |
| Miércoles | Zona 2 o caminata | 45-60 min | 65-75 % FCmax, conversación posible | Volumen, recuperación |
| Jueves | HIIT o intervalos | 25-35 min | 8×2 min al 95 % Wmax | Señal AMPK, mitofagia |
| Viernes | Fuerza compuesta | 45-60 min | Alternado con el martes | Composición, salud metabólica |
| Sábado | Zona 2 largo | 60-90 min | Conversación posible | Volumen para contenido |
| Domingo | Descanso activo | — | Caminar, movilidad | Recuperación |
Tres rasgos del esquema valen explicitar. Primero, dos sesiones por encima de la zona 2, que es donde se fabrica la función mitocondrial. Segundo, dos sesiones de fuerza, que no sustituyen al aeróbico para la mitocondria pero son imprescindibles para la masa muscular. Tercero, la zona 2 aparece dos veces como acumulador de volumen, no como única herramienta. El detalle de protocolos (4×4, 8×2 y variantes) está en el satélite de entrenamiento del cluster VO2max.
Una imagen fija la jerarquía: el ejercicio de endurance rescata el fenotipo progeroide completo en ratones con mutaciones catastróficas en el ADN mitocondrial (Safdar, Tarnopolsky y colaboradores, PNAS 2011) [14]. No existe fármaco que haga eso. No existe suplemento que haga eso. Por eso, en la jerarquía clínica, el ejercicio está en la cúspide y todo lo demás optimiza al margen.
El cambio requiere menos minutos, no más. Si tienes 3-4 horas semanales y las gastas enteras en zona 2, estás optimizando volumen a costa de intensidad. Para la mayoría de adultos no atletas, añadir dos sesiones cortas por encima del umbral láctico produce más adaptación mitocondrial que una hora extra de paseo suave.
Preguntas frecuentes
¿Qué tipo de ejercicio fabrica mitocondrias?
El ejercicio aeróbico de alta intensidad es el que mejor induce biogénesis mitocondrial. En el estudio con biopsia muscular más limpio (Robinson 2017), solo el HIIT y el entrenamiento combinado mejoraron la respiración mitocondrial; el entrenamiento de fuerza aislado no lo hizo. La intensidad por encima del umbral láctico, combinada con suficiente volumen, es la palanca principal.
¿Es mejor el HIIT o la zona 2 para las mitocondrias?
Depende de qué quieras construir y de cuántas horas entrenes. El HIIT es superior para la función mitocondrial y muestra adaptaciones detectables en 2-4 semanas. La zona 2 construye contenido, pero solo con volúmenes altos y sesiones largas. Para quien tiene horas limitadas (3-5 a la semana), la evidencia apoya priorizar trabajo por encima de la zona 2; la zona 2 funciona mejor como acumulador de volumen que como única herramienta.
¿Sirve el entrenamiento de fuerza para las mitocondrias?
Para biogénesis mitocondrial, no de forma comparable al aeróbico intenso. La fuerza activa más la vía mTORC1 (síntesis proteica e hipertrofia) que la AMPK (biogénesis), y en biopsia no mejora la respiración mitocondrial. Eso sí, la fuerza es imprescindible por otras razones: masa muscular, densidad ósea y sensibilidad a la insulina. Fuerza y aeróbico son complementarios, no intercambiables.
¿Cuánta zona 2 hay que hacer?
La zona 2 tiene valor real como trabajo de volumen sostenible y de bajo riesgo, pero no conviene que sea tu única herramienta si buscas salud mitocondrial con horas limitadas. Su señal de PGC-1α solo aparece de forma consistente con sesiones largas, de 60-90 minutos. Un esquema razonable la incluye dos veces por semana como acumulador de volumen, junto con dos sesiones por encima del umbral.
¿Qué construye más mitocondrias, el volumen o la intensidad?
Construyen cosas distintas, no compiten. El volumen total de entrenamiento es el principal motor del contenido mitocondrial (la cantidad de masa mitocondrial), mientras que la intensidad es la que mejora la función respiratoria de cada mitocondria. Para optimizar ambas dimensiones hay que combinar volumen e intensidad, no elegir una.
¿Cuánto tarda el ejercicio en aumentar las mitocondrias?
Las adaptaciones empiezan pronto: la respiración mitocondrial puede subir alrededor de un 22 % tras solo 2 semanas de HIIT en personas no entrenadas, y los cambios de contenido son detectables en 2-4 semanas. El matiz importante es que estas adaptaciones revierten rápido cuando cae el volumen de entrenamiento, así que la constancia es lo que sostiene la ganancia.
¿La metformina interfiere con las adaptaciones del ejercicio?
Sí. Un ensayo doble ciego (Konopka 2019) mostró que la metformina anuló el aumento de VO2max y bloqueó la mejora de respiración mitocondrial inducida por 12 semanas de HIIT en adultos mayores. En personas no diabéticas que entrenan, tomar metformina «por longevidad» puede ser contraproducente para la adaptación mitocondrial que el ejercicio busca construir.
Este artículo es material educativo y de referencia editorial. No constituye consejo médico ni prescripción de entrenamiento individual. Iniciar un programa de HIIT o de trabajo por encima del umbral láctico requiere evaluación cardiovascular previa en personas mayores de 50 años, con factores de riesgo cardiovascular, con patología musculoesquelética significativa o tras periodos prolongados de inactividad. La intensidad y la frecuencia cardiaca máximas deben determinarse de forma individual; la fórmula «220 menos la edad» es una aproximación gruesa con amplio margen de error. El protocolo de la sección 05 es orientativo para el adulto sano entrenado; cualquier persona con historia médica debe ajustarlo con supervisión clínica.
- Memme JM, Sanfrancesco VC, Hood DA. Exercise as mitochondrial medicine: how does the exercise prescription affect mitochondrial adaptations to training? Annu Rev Physiol. 2025;87:23-46. DOI.
- Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell. 1998;92(6):829-839. DOI.
- Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell. 1999;98(1):115-124. DOI.
- Gurd BJ, Menezes ES, Arhen BB, Islam H. Impacts of altered exercise volume, intensity, and duration on the activation of AMPK and CaMKII and increases in PGC-1α mRNA. Semin Cell Dev Biol. 2023;143:17-27. DOI.
- Ristow M, Zarse K, Oberbach A, Klöting N, Birringer M, Kiehntopf M, Stumvoll M, Kahn CR, Blüher M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc Natl Acad Sci USA. 2009;106(21):8665-8670. DOI.
- Robinson MM, Dasari S, Konopka AR, Johnson ML, Manjunatha S, Esponda RR, Carter RE, Lanza IR, Nair KS. Enhanced protein translation underlies improved metabolic and physical adaptations to different exercise training modes in young and old humans. Cell Metab. 2017;25(3):581-592. DOI. PMID 28273480.
- Pengam M, et al. Effects of high-intensity interval training and moderate-intensity continuous training on mitochondrial dynamics in human skeletal muscle. Front Physiol. 2025;16:1554222. DOI.
- Konopka AR, Laurin JL, Schoenberg HM, Reid JJ, Castor WM, Wolff CA, Musci RV, Safairad OD, Linden MA, Biela LM, Bailey SM, Hamilton KL, Miller BF. Metformin inhibits mitochondrial adaptations to aerobic exercise training in older adults. Aging Cell. 2019;18(1):e12880. DOI. PMID 30548390.
- Granata C, Jamnick NA, Bishop DJ. Training-induced changes in mitochondrial content and respiratory function in human skeletal muscle. Sports Med. 2018;48(8):1809-1828. DOI.
- Granata C, Oliveira RS, Little JP, Renner K, Bishop DJ. Training intensity modulates changes in PGC-1α and p53 protein content and mitochondrial respiration, but not markers of mitochondrial content in human skeletal muscle. FASEB J. 2016;30(2):959-970. DOI.
- MacInnis MJ, Gibala MJ. Physiological adaptations to interval training and the role of exercise intensity. J Physiol. 2017;595(9):2915-2930. DOI.
- Granata C, Oliveira RSF, Little JP, Renner K, Bishop DJ. Mitochondrial adaptations to high-volume exercise training are rapidly reversed after a reduction in training volume in human skeletal muscle. FASEB J. 2016;30(10):3413-3423. DOI.
- Storoschuk KL, Moran-MacDonald A, Gibala MJ, Gurd BJ. Much ado about Zone 2: a narrative review assessing the efficacy of Zone 2 training for improving mitochondrial capacity and cardiorespiratory fitness in the general population. Sports Med. 2025;55(7):1611-1624. DOI. PMID 40560504. Conflicto de interés declarado: Gibala con equity en Longevity League Ltd.
- Safdar A, Bourgeois JM, Ogborn DI, Little JP, Hettinga BP, Akhtar M, Thompson JE, Melov S, Mocellin NJ, Kujoth GC, Prolla TA, Tarnopolsky MA. Endurance exercise rescues progeroid aging and induces systemic mitochondrial rejuvenation in mtDNA mutator mice. Proc Natl Acad Sci USA. 2011;108(10):4135-4140. DOI.
- Storoschuk KL, Lessard SJ, Islam H, Gibala MJ, Gurd BJ. High-intensity exercise and mitochondrial biogenesis: current controversies and future research directions. Physiology. 2019;34(1):56-70. DOI.
- Short KR, Bigelow ML, Kahl J, Singh R, Coenen-Schimke J, Raghavakaimal S, Nair KS. Decline in skeletal muscle mitochondrial function with aging in humans. Proc Natl Acad Sci USA. 2005;102(15):5618-5623. DOI. PMID 15800038.