Mitofagia: la limpieza mitocondrial que decide si envejeces bien o te deterioras
La autofagia se ha convertido en el término de moda de la longevidad. El proceso que de verdad decide si tus mitocondrias envejecen bien es más específico —la mitofagia— y la jerarquía de palancas para activarlo no la decide el marketing.
La autofágia se ha convertido en el término de moda de la longevidad. El proceso que de verdad decide si tus mitocondrias envejecen bien es más específico: la mitofagia. Ejercicio, urolitina A y rapamicina la inducen por vías distintas, con magnitudes distintas y con gaps de evidencia que el wellness prefiere no nombrar.
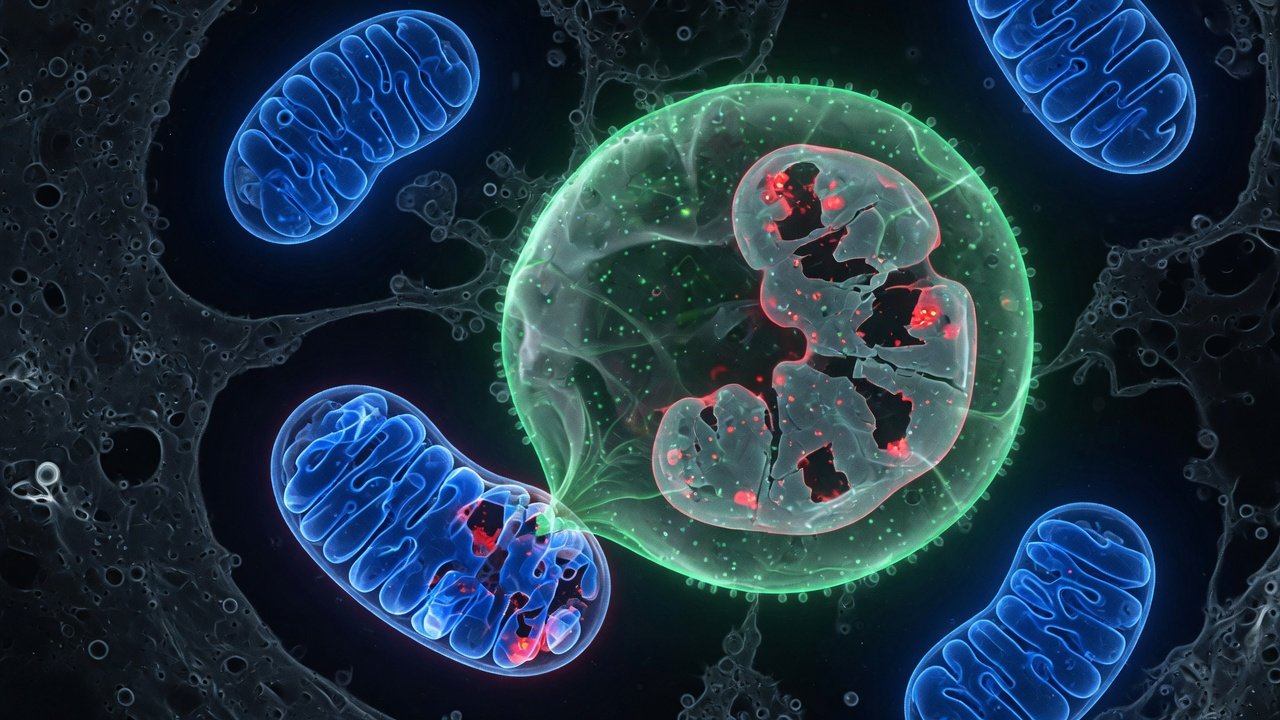
La mitofagia es el proceso por el que una célula identifica, marca y degrada selectivamente sus mitocondrias dañadas. No es autofágia general —no se recicla materia citoplasmática al azar—, es eliminación dirigida del orgánulo defectuoso, una pieza por pieza, sin tocar las sanas. En un sistema biológico que produce energía quemando oxígeno a alta tasa, la mitocondria es a la vez fuente crítica de ATP y fuente crítica de daño oxidativo. La capacidad de retirar la que falla determina la trayectoria de envejecimiento del tejido más que casi cualquier otro marcador aislado.
Este artículo es el hub de KRECE sobre la KW mitofagia. Se centra en músculo, palancas farmacológicas y palancas nutricionales, donde la mayor parte de la evidencia humana existe. Para el ángulo cerebral —mitofagia neuronal, rapamicina y Alzheimer, biogénesis mitocondrial cerebral— existe el satélite paralelo del cluster: mitofagia cerebral y rapamicina en el cerebro envejecido. Las dos piezas se complementan; la línea divisoria es tejido: músculo aquí, cerebro allí.
Mitofagia: el control de calidad selectivo de la mitocondria
Mitofagia en una frase
La mitofagia es la degradación selectiva de mitocondrias dañadas mediante la maquinaria de autofagia. La célula detecta una mitocondria que ha perdido potencial de membrana o sobreproduce ROS, la envuelve en un autofagosoma de doble membrana, y la fusiona con un lisosoma para reciclar sus componentes. Es el sistema de mantenimiento que evita que el tejido acumule orgánulos defectuosos.[1]
Mitofagia vs autofagia general: la diferencia clave
La autofagia general responde a demanda: la célula tiene hambre y recicla lo que pilla. La mitofagia responde a oferta: una mitocondria específica se ha dañado y se «ofrece» para sacrificio. Comparten la maquinaria aguas abajo —autofagosomas, lisosomas— pero las señales de inicio son distintas: privación de nutrientes vs despolarización mitocondrial. Confundir ambas es el error más común cuando el wellness habla de «autofagia» como si fuera un proceso único.
PINK1, Parkin y la vía dominante
La mitocondria dañada pierde potencial transmembrana y eso estabiliza PINK1 en su membrana externa. PINK1 fosforila ubiquitina y recluta a Parkin, una E3 ubiquitin-ligasa, que marca la superficie del orgánulo con cadenas de poliubiquitina. Los adaptadores autofágicos —P62, OPTN, NDP52— reconocen esas marcas y reclutan LC3B, la proteína que define la membrana del autofagosoma. El autofagosoma envuelve la mitocondria y la fusiona con el lisosoma. Los aminoácidos, lípidos y nucleótidos resultantes vuelven al pool celular.[1,3] Existen otras vías por receptor (BNIP3, NIX, FUNDC1, FKBP8) que operan en hipoxia y desarrollo, pero PINK1-Parkin es la dominante en tejido post-mitótico.
Mitochondrial autophagy explained — the short version
In English biomedical literature, the process is called mitochondrial autophagy, mitophagy, or selective autophagy of mitochondria. The three terms describe the same phenomenon: a cell identifies a dysfunctional mitochondrion, tags it with ubiquitin via the PINK1-Parkin axis, and delivers it to the lysosome for degradation. It is the cell’s quality control system for its own energy-producing organelles.
Mitofagia y hallmarks del envejecimiento
La disfunción mitocondrial es uno de los hallmarks del envejecimiento.[11] Dentro de esa disfunción, la caída en la capacidad de mitofagia es el bucle que conecta todo lo demás. Sin reciclaje, las mitocondrias defectuosas se acumulan, generan estrés oxidativo crónico, activan el inflammaging desde la propia mitocondria como hub de señalización, y eventualmente desencadenan apoptosis o senescencia celular. Más contexto en el ancla del cluster.
¿En qué tejidos opera la mitofagia?
La mitofagia es universal: opera en todos los tejidos con mitocondrias, es decir, todos excepto los eritrocitos maduros. Pero la dependencia funcional cambia. Músculo esquelético: mitofagia inducida principalmente por estrés energético (contracción, ayuno), vía dominante AMPK-ULK1. Músculo cardíaco: mitofagia continua, basal alta, mecanismo similar. Cerebro: vía PINK1-Parkin dominante, neurona post-mitótica con dependencia crítica; tratamiento específico en la pieza paralela. Hígado: muy regulada por mTOR y ayuno. Tejido adiposo: regulada por estado nutricional y ejercicio. La mayor parte de la evidencia humana viene de biopsias musculares por razones prácticas: es el tejido más accesible.
Mitofagia vs autofágia, recapitulando. La autofágia responde a demanda energética (la célula tiene hambre y recicla lo que pilla). La mitofagia responde a oferta —una mitocondria específica se ha dañado y se ofrece para ser sacrificada—. Ambas comparten la maquinaria final, pero las señales de inicio son distintas: privación de nutrientes vs despolarización mitocondrial. Cuando un suplemento promete «activa la autofágia», está vendiendo una etiqueta genérica que cubre dos procesos distintos.
Medir mitofagia: el problema de capturar dinámica con fotografías estáticas
Por qué medir mitofagia en humanos sigue siendo casi imposible
En humanos, la mitofagia no se puede medir directamente.[2] Lo que se mide son snapshots moleculares: expresión génica o abundancia de proteínas asociadas (PINK1, Parkin, P62, LC3B, ULK1, FUNDC1) en biopsias musculares, mediante western blot o PCR.[3] Esto da una foto fija del momento de la biopsia, no el flujo real de mitocondrias siendo degradadas. Cuando un ensayo dice «aumentó la mitofagia» en humanos, casi siempre lo que aumentó fue la maquinaria medida en una biopsia, no el proceso.
Los tres modelos animales: MitoQC, MitoKeima, MitoTimer
La medición dinámica directa solo es posible en modelos animales transgénicos con sondas fluorescentes. Hay tres sistemas relevantes, y no son equivalentes:
| Modelo | Mecanismo | Ventaja | Limitación |
|---|---|---|---|
| MitoQC[9] | mCherry-GFP anclado a membrana externa (vía FIS-1). Neutro: verde+rojo (amarillo). Lisosoma ácido: solo rojo. | Cuantificación directa del evento de degradación lisosomal. | Requiere microscopía confocal avanzada. Autofluorescencia tisular interfiere. |
| MitoTimer[21] | Proteína fluorescente temporizada: verde (recién sintetizada) → rojo (madura/estrés oxidativo). | Mide la «edad» global del reteículo mitocondrial. | No mide el evento de mitofagia específico, sino recambio estructural. |
| MitoKeima | Proteína resistente a proteasas lisosomales. Espectro de excitación cambia según pH (citoplasma vs lisosoma). | Señal bimodal muy sensible. | La fijación química destruye el gradiente de pH. Solo tejido fresco o vivo. |
Marcadores indirectos en biopsia muscular
Lo que se mide en humanos en práctica investigadora:
- PINK1 — quinasa estabilizada en mitocondrias dañadas.
- Parkin — ubiquitin-ligasa que marca el orgánulo.
- P62 / SQSTM1 — adaptador autofágico; su acumulación indica bloqueo del flujo.
- LC3B-II — forma lipidada de LC3, marcador de autofagosoma formado.
- ULK1 — quinasa iniciadora; el ratio fosforilado/total indica activación.
- FUNDC1, BNIP3, NIX — receptores alternativos de mitofagia.
Dinámica mitocondrial: fisión y fusión
El otro lado de la ecuación es la maquinaria que controla la dinámica mitocondrial: fusión (unión de mitocondrias para formar redes tubulares) y fisión (separación de fragmentos dañados para que la maquinaria autofágica los capture). Las proteínas relevantes son OPA1[5] y MFN2[6] para fusión; DRP1[7] y FIS1[8] para fisión. Sin fisión previa no hay mitofagia eficiente: la maquinaria del autofagosoma no puede engullir una red mitocondrial entera; solo fragmentos individuales.[4]
How is mitophagy measured? — the short answer
Directly in animal models using fluorescent reporters (MitoQC, MitoKeima, MitoTimer) that distinguish between mitochondria in neutral cytoplasm and acidic lysosomes. Indirectly in humans through Western blot or PCR analysis of muscle biopsies for proteins associated with the mitophagy pathway (PINK1, Parkin, P62, LC3B). The indirect measurement captures pathway machinery, not flux, and is the main reason translating mouse mitophagy data to human clinical outcomes is harder than it looks.
Limitación crítica. Ningún estudio en humanos ha utilizado modelos de reportero dinámico para mitofagia. Toda la evidencia humana descansa sobre marcadores indirectos. Cuando un ensayo clínico dice «aumentó la mitofagia» en humanos, lo que realmente midió fue un incremento en la expresión de genes o proteínas asociadas —no el flujo real—. Esto no invalida los hallazgos, pero obliga a poner una distancia entre lo que se mide y lo que se infiere.[2]
El ejercicio es el activador más potente y el único con medición directa
El ejercicio es la única intervención donde la mitofagia se ha medido directamente mediante modelos de reportero fluorescente en ratón —no solo marcadores indirectos—. Eso lo coloca, en términos de evidencia mecanística, por delante de cualquier suplemento o fármaco.
Ejercicio aeróbico como activador de mitofagia
Una sesión de carrera en cinta de 90 minutos en ratón indujo un 45% de aumento en el ratio MitoQC (verde:rojo), es decir, en el flujo mitofágico medido directamente. Además, usando MitoTimer, el mismo grupo documentó un aumento superior al 400% en los puncta rojos puros (mitocondrias siendo degradadas activamente) a las 6 horas post-ejercicio.[20] La mitofagia volvió a niveles basales a las ~24 horas, lo que establece una regla clínica directa: la mitofagia inducida por ejercicio requiere repetición diaria o casi diaria para mantener el recambio mitocondrial.
HIIT y mitofagia muscular
El entrenamiento por intervalos de alta intensidad activa AMPK con magnitudes superiores al ejercicio aeróbico moderado, y la fosforilación de ULK1 (Ser555) aguas abajo es proporcional. En humanos, biopsias musculares post-HIIT muestran incrementos en LC3B-II y PINK1, marcadores que en biopsia muscular son los más cercanos a una señal de actividad mitofágica. Más contexto en la evidencia real sobre zona 2 y mitocondria: el HIIT y la zona 2 actúan sobre la misma maquinaria pero por vías parcialmente distintas, y el problema clínico no es elegir entre ambas sino combinarlas.
Exercise-induced mitophagy — the molecular pathway
Exercise lowers the ATP:AMP ratio in muscle. AMPK senses this and phosphorylates ULK1 directly at serine 555, initiating autophagosome assembly. Translocation of additional AMPK pools to the mitochondrial outer membrane (Drake 2021) accelerates local mitophagy. Concurrent activation of PGC1α through AMPK-SIRT1 drives mitochondrial biogenesis. The result is a single signal —low energy— that triggers both removal of old mitochondria and synthesis of new ones in coordination.
AMPK-ULK1: la vía molecular completa
La elegancia bioquímica del ejercicio está en que la misma señal —ratio AMP/ATP elevado— activa simultáneamente los dos lados del control de calidad mitocondrial. AMPK fosforila ULK1 (Ser555) iniciando la mitofagia, y en paralelo activa SIRT1 (vía aumento de NAD+) que deacetila a PGC1α permitiéndole entrar al núcleo y disparar el programa de biogénesis mitocondrial inducida por ejercicio (TFAM, NRF1, NRF2 y la cadena respiratoria). Una señal, dos procesos coordinados. Esto es lo que ningún fármaco oral consigue replicar de momento.
Para confirmar la causalidad de AMPK, Laker et al. usaron ratones knockout de la subunidad α2 de AMPK: tras 90 minutos de ejercicio, no se detectó mitofagia alguna.[20] Sin AMPK, no hay mitofagia muscular inducida por ejercicio. La vía no es opcional, es obligatoria.
AMPK mitocondrial localizada (Drake 2021)
El hallazgo de Drake et al. en PNAS (2021) cambió el modelo. Demostraron que AMPK no solo se activa en el citosol —como se asumía—: se transloca físicamente a la membrana mitocondrial tras el ejercicio o el estrés energético.[22] Esta localización mitocondrial de AMPK fosforila ULK1 desde la propia mitocondria, acelerando la mitofagia local. Es un mecanismo de proximidad que explica por qué el ejercicio es tan eficiente: la señal de daño energético, el sensor y la máquina de degradación están en el mismo sitio físico.
Frecuencia óptima: cada 24 horas
El dato de Laker et al. sobre vuelta a basal a las 24 horas tiene una implicación clínica directa: para mantener un flujo mitofágico continuado conviene entrenar diariamente o casi diariamente. La idea popular de «tres sesiones de gimnasio a la semana» es subo´ptima para mitofagia —no para hipertrofia, esa es otra discusión—. El cuerpo no almacena la mitofagia inducida por una sesión; la consume y vuelve a basal.
Implicación clínica. En un estudio humano, 30 minutos de ejercicio de alta intensidad activaron la vía AMPK/ULK1 hasta tal punto que superó por completo el efecto de 16 horas de ayuno; no hubo diferencias en mitofagia entre sujetos que habían ayunado y los que no. El ejercicio maximizó la respuesta por sí solo. Para manipular la mitofagia en consulta, el ejercicio es la palanca más potente y directa —no el ayuno—.
Urolitina A: mitofagia selectiva sin inhibir mTOR
Qué es la urolitina A
La urolitina A (UA) es un posbiótico derivado de los elagitaninos de la granada, fresas, nueces y otros frutos, sintetizado por bacterias intestinales específicas —Gordonibacter urolithinfaciens y Ellagibacter isourolithinifaciens—. Solo el 30-40% de la población posee la microbiota necesaria para producirla endógenamente, lo que justifica la suplementación directa con UA preformada.[23,24]
Mecanismo: mitofagia selectiva sin inhibir mTOR
A diferencia de la rapamicina, la UA no inhibe mTOR. Actúa directamente sobre la mitocondria dañada, induciendo mitofagia selectiva e incrementando biogénesis mitocondrial simultáneamente.[25] Esto significa que no compromete el anabolismo muscular ni la función inmunitaria —un perfil de seguridad radicalmente distinto al de la rapamicina—.
En células musculares, tras 24 horas de exposición a UA, la mitofagia medida con MitoKeima aumentó aproximadamente un 104% respecto al control.[25] En modelos de C. elegans, UA extendió la vida un 45% por mejora de la función mitocondrial.[25]
Evidencia humana en RCTs
Un ensayo controlado aleatorizado de 1.000 mg/día durante 4 meses en adultos de mediana edad reportó mejoras de ~12% en fuerza muscular y ~12% en resistencia.[26] Biopsias musculares mostraron incremento significativo en genes relacionados con mitofagia, ciclo TCA y oxidación de ácidos grasos. En adultos mayores, un ensayo publicado en JAMA Network Open confirmó mejoras en resistencia muscular y salud mitocondrial.[27]
Urolithin A and muscle function — the data, briefly
Three published human RCTs converge on the same picture: 500-1000 mg/day for 4 months produces small but reproducible improvements in muscle endurance and strength in middle-aged and older adults. Muscle biopsies show upregulation of mitophagy and oxidative phosphorylation gene programs. Mitochondrial function biomarkers improve. The effect size is modest —not transformative— but it is the best-documented evidence of a non-prescription compound activating mitochondrial quality control in human muscle.
Limitaciones y gaps de evidencia
Tres puntos honestos. Uno, todos los ensayos clínicos pivotantes de UA han sido financiados por Amazentis/Timeline, la empresa comercializadora. Eso no invalida los datos —los métodos son rigurosos— pero la replicación independiente sigue siendo escasa. Dos, ningún ensayo ha comparado UA frente a ejercicio físico pautado como brazo activo. Es la pregunta clínicamente relevante —¿aporta UA algo que el ejercicio no aporte ya?—, y no hay respuesta directa hoy. Tres, la variabilidad individual en producción endógena de UA es un confusor enorme. Un «respondedor de UA» produce UA endógena desde elagitaninos dietéticos; un no respondedor no. La suplementación con UA preformada equipara a ambos, pero los efectos clínicos reales pueden diferir.[23,24]
UA en cerebro. La aplicación cerebral de UA está mucho menos desarrollada en humanos. Existen datos preclínicos prometedores en modelos animales de Alzheimer y envejecimiento, pero ningún RCT publicado con endpoint cognitivo primario. Amazentis ha anunciado públicamente que la salud cerebral es su próxima área de investigación clínica. Por ahora, UA es una intervención con evidencia muscular sólida y aplicación cerebral pendiente.
Rapamicina: el mejor expediente preclínico del campo y el primer dato humano sobrio
Mecanismo: inhibición mTORC1 → autofagia/mitofagia
La rapamicina inhibe alostéricamente a mTORC1, un complejo que en estado basal frena la maquinaria autofágica fosforilando ULK1 en sitios inhibitorios. Cuando mTORC1 se apaga, ULK1 queda libre para ser fosforilada por AMPK en los sitios activadores, y el autofagosoma arranca. La rapamicina, por tanto, no induce mitofagia directamente: levanta el freno que el estado nutricional impone sobre la maquinaria. Más contexto en el Bio de rapamicina de KRECE.
Datos preclínicos en músculo: McWilliams 2022
El estudio de McWilliams y colaboradores en Cell Metabolism (2022) usa el modelo Deletor —ratones con mutaciones en el ADN mitocondrial que reproducen una enfermedad mitocondrial muscular humana—.[28] El hallazgo central es que la mitofagia muscular en estos animales es heterogénea: hay fibras con mitofagia activa (rescate) y fibras con mitofagia detenida (halting), en patrón de mosaico. La rapamicina restaura el flujo en las fibras con halting. El paper se centra en músculo —no en cerebro envejecido—, y los autores no extrapolan los hallazgos más allá de la enfermedad mitocondrial primaria.
Modelo Deletor ¿= envejecimiento? Es un error común en literatura divulgativa tratar al modelo Deletor como modelo de envejecimiento mitocondrial genérico. Los Deletor llevan mutaciones específicas que aceleran el daño mitocondrial; el envejecimiento natural es más lento y multifactorial. La extrapolación «si rapamicina rescata Deletor, rescata envejecimiento» es un salto que los datos del paper no sostienen. KRECE evita esa simplificación.
Mitofagia cerebral con rapamicina: el ángulo Brain
La rapamicina activa mitofagia en cerebro envejecido y modelos de Alzheimer en ratones, con un mecanismo dual interesante: elimina mitocondrias dañadas y simultáneamente aumenta síntesis proteica mitocondrial en cerebro envejecido. La narrativa preclínica está bien construida y descansa en Wang 2021 (modelo APP/PS1) y Reid 2020 (UM-HET3) —con matices sex-specific que conviene leer en detalle—. Pero la primera traducción a humanos —el Phase 1 piloto Gonzales 2025 en MCI/Alzheimer temprano— mostró que la rapamicina a 1 mg/día durante 8 semanas no es detectable en LCR y no produjo cambios cognitivos.
El tratamiento exhaustivo del ángulo cerebral —mTORC1 neuronal vs muscular, biogénesis mitocondrial cerebral, hipótesis Kaeberlein-Galvan, Gonzales 2025, nuevo ensayo UT Health San Antonio de marzo 2026— vive en el satélite paralelo del cluster: mitofagia cerebral, rapamicina y Alzheimer en el cerebro envejecido. Aquí la pieza cierra el círculo con la mirada muscular y de longevidad sistémica.
ITP y extensión de vida: el expediente más sólido en geroprotección
El Interventions Testing Program (ITP) del NIA/NIH —el programa de longevidad preclínica más riguroso del mundo, con tres colonias independientes— posiciona a la rapamicina como el compuesto con mayor extensión de vida documentada:
| Dosis (ppm) | Cohorte | Inicio | Machos | Hembras |
|---|---|---|---|---|
| 14 | C2005/C2006 | 9 meses | +8-10% | +17-22% |
| 42 | C2009 | 9 meses | +22% | +28% |
| 14 | C2009 | 20 meses | +20% | +13% |
| 42 intermitente | C2015 | 9 meses | +8-10% | +8-10% |
| 14.7 + Acarbosa 1000 | C2017 | 9 meses | +37% | Pendiente |
Dos datos críticos: el inicio tardío (20 meses, equivalente a 60-65 años humanos) funcionó casi tan bien como el inicio temprano. Y la combinación rapamicina + acarbosa logró el mayor incremento de lifespan jamás documentado en el ITP: +37% en machos. Más contexto y ensayos clínicos en humanos en el blog editorial sobre rapamicina, ITP y PEARL.
Phase 1 humano (Gonzales 2025): la actualización 2026
En mayo de 2025, Gonzales y colaboradores publicaron en Communications Medicine el primer Phase 1 piloto de rapamicina en pacientes con MCI o Alzheimer temprano: 10 participantes, 1 mg/día durante 8 semanas.[33] Los hallazgos:
- Rapamicina no detectable en LCR antes ni después del tratamiento (endpoint primario).
- Sin cambios cognitivos significativos.
- HbA1c y presión arterial sistólica aumentaron.
- p-tau-181, GFAP y NfL en LCR aumentaron significativamente (interpretación ambigua: empeoramiento vs aumento de turnover proteico).
- 20 eventos adversos, mayoría leves.
El dato no anula la hipótesis geroprotectora —la rapamicina sigue siendo el compuesto con mejor expediente en ITP—, pero sí obliga a recalibrar el discurso sobre rapamicina cerebral en humanos. Más detalle en la pieza paralela sobre mitofagia cerebral, rapamicina y Alzheimer.
Rapamycin and mitophagy — the long view
Rapamycin remains the gold-standard preclinical geroprotector in the ITP database: consistently extends lifespan across sexes, doses and starting ages, including late-life initiation. Its mechanism on mitophagy is indirect —mTORC1 inhibition releases ULK1 to initiate autophagosome assembly— rather than direct mitochondrial action. Human clinical translation for longevity is in early phases: Mannick 2014 demonstrated safe immune benefits with low intermittent doses; Gonzales 2025 demonstrated absence of CSF penetration at 1 mg/day in dementia patients; the new UT Health San Antonio NIA-funded trial (n=84) is expected to inform dosing in healthy older adults over the next several years.
Todo N1 en músculo, N4 inicial en humanos. La rapamicina tiene el expediente preclínico más robusto del mundo, pero los ensayos clínicos humanos para longevidad o cognición están en fases muy tempranas. Las dosis geroprotéctoras propuestas (5-6 mg/semana, pauta intermitente) no están validadas por ningún ECA con endpoint duro de longevidad o salud cognitiva. Más contexto y limitaciones en la guía de rapamicina.
Metformina: el fármaco que bloquea lo que debería potenciar
Mecanismo: Complejo I → AMPK → mitofagia
La metformina inhibe parcialmente el Complejo I de la cadena respiratoria mitocondrial, altera el ratio AMP:ATP y activa AMPK secundariamente. En teoría, debería inducir mitofagia. Drake et al. (2021) demostraron que efectivamente induce localización mitocondrial de AMPK, igual que el ejercicio.[22] Hasta aquí, la lógica mecánica funciona.
Fracaso en el ITP
Metformina en monoterapia (1000 ppm, cohorte C2011, inicio 9 meses) no extendió la vida en ratones sanos: +7% en machos (p=0.35), 0% en hembras (p=0.79).[ITP] La molécula que millones de personas toman como «píldora de longevidad» fracasó en el programa más riguroso del mundo. La sinergia solo aparece al combinarla con rapamicina: MetRapa (metformina 1000 ppm + rapamicina 14 ppm) logró +23% en machos y +23% en hembras. La sinergia es real en machos (donde rapamicina sola daba 8-10%), pero en hembras es redundante (rapamicina sola ya daba 17-22%).
Antagonismo con ejercicio (Peterson et al.): el blunting effect
El dato más relevante para la práctica clínica: la metformina bloquea las adaptaciones mitocondriales del ejercicio. Un ensayo controlado aleatorizado en adultos mayores comparó ejercicio + metformina vs ejercicio + placebo durante 12 semanas. El grupo de metformina experimentó una atenuación severa: más del 50% de reducción en la mejora de VO2 máx, bloqueo del incremento en respiración mitocondrial muscular, y anulación de la mejora en sensibilidad a la insulina que normalmente induce el ejercicio.
El mecanismo es claro: la metformina inhibe el Complejo I, que es exactamente la señal que el ejercicio utiliza para disparar la biogénesis mitocondrial y la mitofagia. Tomar metformina antes de entrenar es como pisar el acelerador y el freno a la vez.
Posición clínica. Para un paciente diabético sedentario, la metformina tiene sentido metabólico. Para un paciente que entrena regularmente y busca optimización mitocondrial, la metformina es contraproducente. La lógica de «más intervenciones = mejor» se estrella contra la bioquímica: cuando dos estímulos compiten por la misma vía pero en direcciones opuestas, el resultado no es sinergia sino cancelación.
Espermidina, restricción calórica y precursores de NAD+
Espermidina: la palanca dietética con señal humana
La espermidina es una poliamina endógena que induce autofagia y mitofagia mediante inhibición de la acetiltransferasa EP300.[15] El estudio de Eisenberg et al. (Nature Medicine, 2016) demostró extensión de vida en ratones con un mecanismo específicamente dependiente de mitofagia cardíaca —no autofagia general—. En ratones con los genes de autofagia anulados (Atg5 knockout), la espermidina no tuvo ningún efecto cardioprotector, demostrando que su poder depende al 100% de la capacidad de la célula para reciclar mitocondrias dañadas.[15]
En humanos, el ensayo SmartAge (Wirth et al.) en mayores con quejas subjetivas de memoria documentó mejoras cognitivas tras 3 meses de espermidina derivada de germen de trigo (~1.2 mg/día). Más detalle en el satélite dedicado: espermidina, autofagia y protocolos.
Restricción calórica
La restricción calórica activa AMPK, suprime mTOR e induce mitofagia. Pero la respuesta es extraordinariamente heterogénea: en modelos animales crónicos, solo un tercio de los ratones se benefició; otro tercio tuvo respuestas neutras; y el último tercio experimentó respuestas abiertamente perjudiciales, dependiendo del background genético. No es una intervención universal. En humanos, el ensayo CALERIE-2 (n=218, restricción calórica del 25% durante 2 años) documentó mejoras en marcadores de envejecimiento biológico medidos por reloj epigenético.
Precursores de NAD+ (NR y NMN)
El Nicotinamída Ribósido (NR) y el NMN son precursores de NAD+, cofactor esencial para la cadena respiratoria y las sirtuinas. En ratones, las dosis han sido masivas (400-500 mg/kg) con resultados impresionantes en biogénesis mitocondrial. En humanos, la evidencia ha fracasado consistentemente. Un estudio en 40 hombres obesos con 2.000 mg de NR durante 12 semanas no encontró ninguna mejora en sensibilidad a la insulina, daño oxidativo ni función mitocondrial. Biopsias musculares en ensayos de 4-6 semanas mostraron que la suplementación oral con NR ni siquiera logró incrementar los niveles intramusculares de NAD+. La desconexión traslacional es total: lo que funciona a 500 mg/kg en ratones no funciona a 2.000 mg totales en humanos.
Antioxidantes: la intervención que destruye lo que intenta proteger. Las megadosis de vitaminas C y E bloquean los picos transitorios de ROS que las mitocondrias necesitan como señal de alarma para disparar la biogénesis y la mitofagia. Los antioxidantes sintéticos no solo no mejoran la salud mitocondrial —la sabotean activamente al eliminar la señal de hormesis—.
Comparativa de palancas y los gaps que el wellness no nombra
Tabla comparativa: magnitud, método, modelo, nivel
| Intervención | Magnitud | Método | Modelo | Nivel |
|---|---|---|---|---|
| Ejercicio (90 min carrera) | +45% (MitoQC), >400% puncta rojos (MitoTimer) | Reportero fluorescente directo | Ratón | N1 directo |
| Urolitina A (24h, células) | ~104% (MitoKeima) | Reportero fluorescente directo | Células musculares | N0 directo |
| Urolitina A (1000 mg/d, 4 meses) | +12% fuerza, +12% resistencia | Endpoints funcionales + expresión génica | Humanos (mediana edad) | N4 |
| Rapamicina (modelo Deletor) | Restauración del flujo en fibras con halting | MitoQC + western blot | Ratón (enf. mitocondrial muscular) | N1 (modelo específico) |
| Rapamicina (modelos AD) | Aumenta mitofagia hipocampal (Parkin-mediada) | Co-localización TOM20/LC3B | Ratón APP/PS1 | N1 |
| Rapamicina (humano MCI/AD) | No detectable en LCR, sin cambio cognitivo a 8 sem | Espectrometría de masas LCR | Humanos n=10 (Gonzales 2025) | N3 (piloto) |
| Espermidina (Eisenberg 2016) | Extensión lifespan, cardioprotección | Dependiente de Atg5 (mitofagia) | Ratón | N1 |
| Espermidina (SmartAge) | Mejora cognitiva en mayores con queja de memoria | Tests cognitivos vs placebo | Humanos n=∼30 | N4 |
| NR/NMN (2000 mg, 12 sem) | Sin efecto mitocondrial detectable | Biopsias musculares | Humanos (obesos) | N4 negativo |
Los tres gaps que nadie quiere nombrar
Gap 1 — Ejercicio + Urolitina A en humanos. No existe ningún ECA que combine ambas intervenciones con brazo activo de ejercicio. No sabemos si UA aporta algo adicional a un paciente que ya entrena, o si sus beneficios se limitan a rescatar la maquinaria oxidativa de individuos sedentarios. Es exactamente la pregunta que más importa clínicamente y la que menos se está respondiendo.
Gap 2 — Ejercicio + Rapamicina en humanos. No hay datos —ni en humanos ni en roedores adecuadamente potenciados— sobre si las dosis geroprotéctoras de rapamicina bloquean la hipertrofia muscular o la reparación tisular post-entrenamiento. Con metformina sabemos que sí (Peterson et al.). Con rapamicina, la pregunta sigue abierta.
Gap 3 — Combinaciones triple. La teoría de que dieta cetogénica + metformina + ayuno constituiría una «terapia metabólica hiperpotente» por convergencia en AMPK y mTOR no tiene un solo estudio clínico que la respalde. Es empirismo mecanístico puro vendido como protocolo.
Para profundizar la jerarquía de palancas
Cada intervención tiene su nicho. Una visión integrada que ordena cuándo y por qué cada palanca encaja en un plan de longevidad mitocondrial está en la jerarquía KRECE de palancas mitocondriales con evidencia. Si esta pieza es el hub mitofagia, esa es el hub funcional del cluster.
La mitofagia no es un indicador más — es el indicador
Preguntas frecuentes sobre mitofagia
¿Qué es la mitofagia en una frase?
La mitofagia es la degradación selectiva de mitocondrias dañadas dentro de una célula mediante autofagia: el sistema de mantenimiento que evita que las mitocondrias defectuosas se acumulen y generen estrés oxidativo crónico.
¿Cuál es la diferencia entre mitofagia y autofagia?
La autofagia es el proceso general por el que la célula recicla componentes citoplasmáticos en respuesta a privación de nutrientes. La mitofagia es una forma selectiva que elige específicamente mitocondrias dañadas. Comparten la maquinaria final (autofagosoma + lisosoma), pero las señales de inicio son distintas.
¿Cómo se activa la mitofagia?
Por dos rutas. Ruta AMPK: estrés energético (ejercicio, ayuno) que baja el ratio ATP/AMP, AMPK fosforila ULK1, arranca autofagosoma. Ruta mTOR: privación de nutrientes o inhibición farmacológica (rapamicina) que apaga mTORC1, liberando ULK1 del freno inhibitorio. El ejercicio activa ambas rutas a la vez.
¿Cómo se activa la mitofagia de forma natural?
Ejercicio aeróbico o HIIT (la palanca más potente y mejor documentada), ayuno prolongado (24-72 h), sueño de calidad (sistema glinfático nocturno), restricción calórica cíclica, y dieta rica en alimentos con elagitaninos (granada, fresas, nueces) si tu microbioma produce urolitina A endógena.
¿Cuáles son los síntomas de mitofagia deficiente?
La mitofagia no se mide clínicamente, no hay síntomas específicos. Lo que sí correlaciona con disfunción mitocondrial generalizada en humanos: fatiga inexplicable, intolerancia al ejercicio, pérdida de fuerza desproporcionada para la edad, deterioro cognitivo en envejecimiento, debilidad muscular post-COVID. Son cuadros multifactoriales; la mitofagia es solo una pieza del rompecabezas.
¿Cuánto tiempo de ayuno se necesita para activar mitofagia?
En humanos, los cambios moleculares medibles (marcadores autofágicos en leucocitos circulantes) aparecen a partir de 16-24 horas de ayuno y se intensifican con duraciones más largas. Pero un estudio clásico mostró que 30 minutos de ejercicio de alta intensidad superan el efecto de 16 horas de ayuno sobre mitofagia muscular medida. El ejercicio es palanca más eficiente que el ayuno para esto.
¿Qué tipo de ejercicio activa más la mitofagia?
El ejercicio aeróbico continuo de intensidad moderada-alta (50-70% VO2 máx, 60-90 minutos) y el HIIT (intervalos al 85-95% durante 4-6 minutos repetidos). Ambos activan AMPK con potencia. El entrenamiento de fuerza también activa mitofagia muscular, pero la magnitud documentada es menor que con aeróbico/HIIT.
¿Cuál es la dosis efectiva de urolitina A?
Los ensayos clínicos pivotantes usan 500 mg/día o 1.000 mg/día durante 4 meses como mínimo. La dosis de 500 mg/día es la más frecuentemente comercializada (Mitopure® de Timeline). Hay que recordar que se necesita continuidad: efectos funcionales aparecen tras 8-12 semanas, no en días.
¿Qué es mejor para la mitofagia: ejercicio, ayuno o suplementos?
Ejercicio, por orden de evidencia humana y magnitud documentada. La pirámide clínica KRECE: ejercicio > urolitina A > espermidina > ayuno periódico > rapamicina experimental. Construir desde abajo: empezar por ejercicio diario o casi diario, suplementar después, considerar farmacología solo bajo supervisión.
¿La rapamicina activa la mitofagia en humanos?
Activa la maquinaria autofágica indirectamente al inhibir mTORC1, pero la evidencia humana de mitofagia tisular real es escasa. En el primer Phase 1 piloto en MCI/Alzheimer (Gonzales 2025, n=10, 1 mg/día, 8 semanas), la rapamicina no fue detectable en LCR y no produjo cambios cognitivos. Sigue siendo intervención experimental supervisada.
¿Es lo mismo mitofagia en músculo que mitofagia en cerebro?
No del todo. El músculo activa mitofagia principalmente vía AMPK-ULK1 en respuesta a estrés energético agudo. La neurona post-mitótica usa principalmente la vía PINK1-Parkin como mecanismo basal continuo. La evidencia humana de mitofagia muscular existe vía biopsia; la mitofagia cerebral directa en humanos vivos sigue siendo imposible de medir. El ángulo cerebral, en detalle, en la pieza paralela sobre mitofagia cerebral y rapamicina en Alzheimer.
¿Qué relación hay entre mitofagia y sarcopenia?
La sarcopenia —pérdida de masa y fuerza muscular con la edad— correlaciona con disfunción mitocondrial muscular: menos mitocondrias funcionales por fibra, peor respiración, más ROS. Mejorar mitofagia muscular es una de las intervenciones plausibles para frenar sarcopenia, junto con entrenamiento de fuerza y ingesta proteica adecuada. Los ensayos de urolitina A en mayores muestran mejoras de resistencia muscular consistentes con esto.
¿Cómo se mide la mitofagia en la práctica clínica?
No se mide en práctica clínica humana estándar. En investigación se usan biomarcadores indirectos en biopsia muscular (PINK1, Parkin, P62, LC3B-II, ULK1 fosforilada) mediante western blot o PCR. La medición dinámica directa requiere modelos animales transgénicos con sondas fluorescentes (MitoQC, MitoKeima, MitoTimer).
¿La metformina activa la mitofagia?
Sí en mecanismo (inhibe Complejo I → sube AMP/ATP → activa AMPK → mitofagia vía ULK1), pero fracasó en el ITP en monoterapia (no extendió vida en ratones sanos) y bloquea las adaptaciones mitocondriales del ejercicio en humanos. Es contraproducente para quien entrena. Tiene sentido en paciente diabético sedentario.
¿Es seguro combinar urolitina A con rapamicina?
No hay datos clínicos sobre la combinación. En teoría actuan por mecanismos distintos (UA directo sobre mitocondria, rapamicina vía mTORC1) y no se canibalizan. En práctica, la rapamicina es prescripción médica supervisada; cualquier combinación debe pasar por criterio clínico, no por lógica suplementaria autodirigida.
¿Cómo afecta el sueño a la mitofagia?
El sueño NREM profundo activa el sistema glinfático, que drena residuos metabólicos del cerebro. Aunque eso no es estrictamente mitofagia (es drenaje extracelular), es complementario: la mitofagia limpia dentro de la célula, el glinfático drena fuera. Sueño de mala calidad afecta directamente la capacidad de reciclaje celular global.
How does exercise induce mitophagy?
Exercise lowers muscle ATP and raises AMP. AMPK senses this and phosphorylates ULK1 at serine 555, initiating autophagosome formation. A portion of AMPK translocates to the mitochondrial outer membrane (Drake 2021), accelerating local mitophagy. In parallel, AMPK activates PGC1α through SIRT1, triggering biogenesis of new mitochondria. The result: one signal, two coordinated processes that remove damaged mitochondria and build new ones in tandem.
How to measure mitophagy in humans?
Directly, you can’t. Indirectly, through Western blot or PCR analysis of muscle biopsies for proteins associated with the mitophagy pathway: PINK1, Parkin, P62/SQSTM1, LC3B-II, phosphorylated ULK1. These markers capture pathway machinery rather than dynamic flux, which is why translating animal data with fluorescent reporters (MitoQC, MitoKeima) to human clinical reality requires careful interpretation.
What is the best mitophagy supplement?
By human RCT evidence: urolithin A (500-1000 mg/day for 4 months) is the best-documented supplement showing functional improvements in muscle strength and endurance. It induces mitophagy without inhibiting mTOR. Spermidine has the SmartAge trial showing cognitive benefit in older adults with subjective memory complaints. NR and NMN have failed consistently in human trials for mitochondrial endpoints despite strong animal data.
¿Mitofagia y autofágia inducidas por ayuno extienden la vida?
En modelos animales sí, con magnitudes significativas (45-50% de extensión con días alternos en ratones). En humanos no hay un ECA con duración suficiente y endpoint duro de longevidad. Los datos humanos disponibles son sobre biomarcadores intermedios (CALERIE-2 mostró mejoras en reloj epigenético). La traslación directa ratón-humano del «tiempo de ayuno» sigue discutida.
¿Mitofagia y cerebro envejecido — resumen?
La mitofagia cerebral es crítica para el envejecimiento cognitivo saludable. La rapamicina la activa en modelos animales, con mejora cognitiva en modelos Alzheimer (Wang 2021, APP/PS1). En humanos con MCI/Alzheimer temprano, el Phase 1 piloto Gonzales 2025 mostró ausencia de penetración en LCR a dosis tolerables. El nuevo ensayo NIA-funded de n=84 en UT Health San Antonio (marzo 2026) puede aportar datos a 2-4 años vista. Todo el ángulo cerebral, en detalle, en la pieza paralela del cluster.
Este artículo es contenido editorial de KRECE. No sustituye al criterio médico individualizado. Las decisiones sobre suplementación con urolitina A, espermidina, NR/NMN, o sobre uso de rapamicina/sirolimus/everolimus o metformina, corresponden al médico tratante, que conoce la historia clínica completa del paciente y la legislación aplicable. La rapamicina para indicaciones de longevidad es prescripción off-label en España, UE y EE. UU. y no se debe iniciar sin supervisión clínica. Los datos cuantitativos citados en este artículo provienen de papers verificados con la fuente primaria; la interpretación editorial es de KRECE y puede actualizarse con nuevas evidencias.
- Youle RJ, Narendra DP. Mechanisms of mitophagy. Nat Rev Mol Cell Biol. 2011;12(1):9-14. doi:10.1038/nrm3028.
- Hamacher-Brady A, Brady NR. Mitophagy programs: mechanisms and physiological implications of mitochondrial targeting by autophagy. Cell Mol Life Sci. 2016;73(4):775-795.
- Ashrafi G, Schwarz TL. The pathways of mitophagy for quality control and clearance of mitochondria. Cell Death Differ. 2013;20(1):31-42. PMID: 22743996.
- Liu YJ, McIntyre RL, Janssens GE, Houtkooper RH. Mitochondrial fission and fusion: a dynamic role in aging and potential target for age-related disease. Mech Ageing Dev. 2020;186:111212.
- Tezze C, Romanello V, Desbats MA, et al. Age-associated loss of OPA1 in muscle impacts muscle mass, metabolic homeostasis, systemic inflammation, and epithelial senescence. Cell Metab. 2017;25(6):1374-1389.
- Bell MB, Bush Z, McGinnis GR, Rowe GC. Adult skeletal muscle deletion of Mitofusin 1 and 2 impedes exercise performance and training capacity. J Appl Physiol. 2019;126(2):341-353.
- Favaro G, Romanello V, Varanita T, et al. DRP1-mediated mitochondrial shape controls calcium homeostasis and muscle mass. Nat Commun. 2019;10(1):2576.
- Losón OC, Song Z, Chen H, Chan DC. Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol Biol Cell. 2013;24(5):659-667.
- McWilliams TG, Prescott AR, Allen GFG, et al. Mito-QC illuminates mitophagy and mitochondrial architecture in vivo. J Cell Biol. 2016;214(3):333-345.
- McWilliams TG, Ganley IG. Investigating mitophagy and mitochondrial morphology in vivo using mito-QC. Methods Mol Biol. 2019;1880:621-642.
- Schmauck-Medina T, Molière A, Lautrup S, et al. New hallmarks of ageing: a 2022 Copenhagen ageing meeting summary. Aging (Albany NY). 2022;14(16):6829-6839.
- Sun N, Youle RJ, Finkel T. The mitochondrial basis of aging. Mol Cell. 2016;61(5):654-666. PMID: 26942670.
- Plummer JD, Johnson JE. Extension of cellular lifespan by methionine restriction involves alterations in central carbon metabolism and is mitophagy-dependent. Front Cell Dev Biol. 2019;7:301.
- Palikaras K, Lionaki E, Tavernarakis N. Coordination of mitophagy and mitochondrial biogenesis during ageing in C. elegans. Nature. 2015;521(7553):525-528. PMID: 25896323.
- Eisenberg T, Abdellatif M, Schroeder S, et al. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat Med. 2016;22(12):1428-1438. PMID: 27841876.
- Kerr JS, Adriaanse BA, Greig NH, et al. Mitophagy and Alzheimer’s disease: cellular and molecular mechanisms. Trends Neurosci. 2017;40(3):151-166. PMID: 28190529.
- Vásquez-Trincado C, García-Carvajal I, Pennanen C, et al. Mitochondrial dynamics, mitophagy and cardiovascular disease. J Physiol. 2016;594(3):509-525.
- Ringholm S, Gudiksen A, Frey Halling J, et al. Impact of aging and lifelong exercise training on mitochondrial function and network connectivity in human skeletal muscle. J Gerontol A Biol Sci Med Sci. 2022;77(2):373-383.
- Granata C, Oliveira RSF, Little JP, et al. Mitochondrial adaptations to high-volume exercise training are rapidly reversed after a reduction in training volume in human skeletal muscle. FASEB J. 2016;30(10):3413-3423.
- Laker RC, Drake JC, Wilson RJ, et al. Ampk phosphorylation of Ulk1 is required for targeting of mitochondria to lysosomes in exercise-induced mitophagy. Nat Commun. 2017;8:548. PMID: 28916822.
- Laker RC, Xu P, Ryall KA, et al. A novel MitoTimer reporter gene for mitochondrial content, structure, stress, and damage in vivo. J Biol Chem. 2014;289(17):12005-12015.
- Drake JC, Wilson RJ, Laker RC, et al. Mitochondria-localized AMPK responds to local energetics and contributes to exercise and energetic stress-induced mitophagy. Proc Natl Acad Sci USA. 2021;118(37):e2025932118. PMID: 34504024.
- D’Amico D, Andreux PA, Valdés P, Singh A, Rinsch C, Auwerx J. Impact of the natural compound urolithin A on health, disease, and aging. Trends Mol Med. 2021;27(7):687-699.
- Singh A, D’Amico D, Andreux PA, et al. Direct supplementation with urolithin A overcomes limitations of dietary exposure and gut microbiome variability in healthy adults to achieve consistent levels across the population. Eur J Clin Nutr. 2022;76(2):297-308.
- Ryu D, Mouchiroud L, Andreux PA, et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat Med. 2016;22(8):879-888. PMID: 27400265.
- Singh A, D’Amico D, Andreux PA, et al. Urolithin A improves muscle strength, exercise performance, and biomarkers of mitochondrial health in middle-aged adults: the randomized, double-blind, placebo-controlled ATLAS trial. Cell Rep Med. 2022;3(5):100633.
- Liu S, D’Amico D, Shankland E, et al. Effect of urolithin A supplementation on muscle endurance and mitochondrial health in older adults: a randomized clinical trial. JAMA Netw Open. 2022;5(1):e2144279.
- Mito T, Vincent AE, Faitg J, et al. Mosaic dysfunction of mitophagy in mitochondrial muscle disease. Cell Metab. 2022;34(2):197-208.e5. doi:10.1016/j.cmet.2021.12.017. PMID: 35030325.
- Ubaida-Mohien C, Spendiff S, Lyashkov A, et al. Unbiased proteomics, histochemistry, and mitochondrial DNA copy number reveal better mitochondrial health in muscle of high-functioning octogenarians. eLife. 2022;11:e74335.
- Kaeberlein M, Galvan V. Rapamycin and Alzheimer’s disease: time for a clinical trial? Sci Transl Med. 2019;11(476):eaar4289. doi:10.1126/scitranslmed.aar4289. PMID: 30674654.
- Reid JJ, Linden MA, Peelor FF, Miller RA, Hamilton KL, Miller BF. Brain protein synthesis rates in the UM-HET3 mouse following treatment with rapamycin or rapamycin with metformin. J Gerontol A Biol Sci Med Sci. 2020;75(1):40-49. doi:10.1093/gerona/glz069. PMID: 30864661.
- Wang H, Fu J, Xu X, Yang Z, Zhang T. Rapamycin activates mitophagy and alleviates cognitive and synaptic plasticity deficits in a mouse model of Alzheimer’s disease. J Gerontol A Biol Sci Med Sci. 2021;76(10):1707-1713. doi:10.1093/gerona/glab142. PMID: 34003967.
- Gonzales MM, Garbarino VR, Kautz TF, et al. Rapamycin treatment for Alzheimer’s disease and related dementias: a pilot phase 1 clinical trial. Commun Med (Lond). 2025;5:189. doi:10.1038/s43856-025-00904-9. PMID: 40394335. PMCID: PMC12092812.
- Mannick JB, Del Giudice G, Lattanzi M, et al. mTOR inhibition improves immune function in the elderly. Sci Transl Med. 2014;6(268):268ra179. doi:10.1126/scitranslmed.3009892. PMID: 25540326.
- Wirth M, Benson G, Schwarz C, et al. The effect of spermidine on memory performance in older adults at risk for dementia: a randomized controlled trial. Cortex. 2018;109:181-188. (SmartAge trial).
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: an expanding universe. Cell. 2023;186(2):243-278. doi:10.1016/j.cell.2022.11.001.